
Current insights into the role of uterine natural killer cells in human reproduction
Authors:
Pavel Otevřel
![]()
Authors‘ workplace:
Reprofit International s. r. o., Brno
Published in:
Ceska Gynekol 2026; 91(3): 226-231
Category:
doi:
https://doi.org/10.48095/cccg2026226
Overview
Human reproductive efficiency is relatively low compared with that of other mammals, a phenomenon that may be linked to evolutionary factors. One of the principal biological contributors is the high incidence of aneuploidy in oocytes and embryos, a risk that increases markedly with advancing maternal age. Successful implantation requires not only a genetically normal embryo, but also a receptive endometrium and a functional feto-maternal dialogue. Nevertheless, even after the transfer of a euploid embryo, implantation occurs in only 60–70% of cases, underscoring the importance of the uterine environment for successful embryo attachment. A key element in the implantation process is the immunological balance at the feto-maternal interface. Uterine natural killer (uNK) cells constitute the dominant lymphocyte population within the decidua and play essential roles in implantation, placentation, and fetal immunotolerance. Their function is mediated, among other mechanisms, by interactions with trophoblast cells through killer-cell immunoglobulin-like (KIR) receptors and human leukocyte antigen C (HLA-C) antigens. Imbalances in these interactions can contribute to pregnancy pathologies such as implantation failure, miscarriage, preeclampsia, and fetal growth restriction. Although current diagnostic methods enable assessment of immunological conditions in reproductive tissues, interpreting these findings in the context of fertility or infertility remains challenging. Emerging research directions include the application of molecular genetics and detailed characterization of distinct NK cell subpopulations. Despite recent advances, therapeutic modulation of immunological mechanisms in reproduction remains limited and warrants further investigation.
Keywords:
implantation – uterine NK cells – pregnancy loss – KIR receptors – HLA-C
Introduction
The success rate of human reproduction is low compared to other mammals, which may have evolutionary causes [1]. One of the main biological factors contributing to the low success of embryo implantation is the high incidence of oocyte and consecutively embryo aneuploidy [2]. The risk of oocyte aneuploidy is directly related to maternal age [3]. Successful implantation requires a genetically normal, so-called euploid embryo, a receptive endometrium, and an intact feto-maternal dialogue; meaning properly functioning processes involved in implantation. Only under these conditions can sufficient chances for implantation be ensured. It is well known, particularly due to the widespread use of preimplantation genetic testing of embryos, that even after transferring a euploid embryo, implantation occurs in only 60–70% of cases [4], and a significant proportion of pregnancies end in loss of a euploid embryo [5]. Therefore, research naturally focuses on identifying causes of implantation failure and pregnancy pathologies within the endometrium [6]. One possible cause of implantation failure is immunological imbalance, leading to unsuccessful implantation, early pregnancy loss, or complications later in pregnancy (such as intrauterine growth restriction or preeclampsia). The presumed mechanism behind these pathologies is insufficient or, conversely, excessive invasion of the trophoblast into the endometrium. The term “feto-maternal interface” is commonly used for the area where this invasion occurs, and “implantation window” refers to the time period during which implantation takes place. In humans, the implantation window opens during a specific phase of the menstrual cycle. Only in humans and a few higher primates does the endometrium undergo spontaneous cyclic decidualization [2], regardless of whether the conceptus is present in the woman’s body. The best-studied immune cells subpopulation involved in these processes of implantation and embryonic development are cells known as uterine or decidual natural killer cells (uNK, dNK).
Immune system of the endometrium
The human endometrium represents a highly dynamic system that undergoes periodic structural and functional changes under the influence of steroid hormones. Upon embryo implantation, the endometrium differentiates into the decidua, whose primary roles include histotrophic nourishment of the embryo and subsequent support of placentation while maintaining immunotolerance toward the semi-allogeneic conceptus. A critical component in implantation and immune adaptation is the embryonic tissue known as extravillous trophoblast (EVT). EVT originates at the periphery of the villous trophoblast and invades deeply into the decidua and myometrium. During this process, EVT mediates the remodelling of spiral arteries into high-capacitance, low-resistance vessels, ensuring adequate maternal blood supply to the placenta.
Throughout its invasion into maternal tissue, EVT interacts with components of the maternal immune system. Antigens expressed on the EVT surface serve as ligands for receptors on maternal immune cells, initiating complex signaling cascades.
The lymphocyte population within the endometrium comprises four major cell types: macrophages, T-helper lymphocytes (marked as CD3+CD4+), cytotoxic T-lymphocytes (CD3+CD8+), and NK cells (CD3–CD56+). Their relative abundance fluctuates across the menstrual cycle, decidualization, and pregnancy.
NK cells belong to the family of innate lymphoid cells (ILCs), which function as a critical link between innate and adaptive immunity [7]. Sharing a common progenitor with B - and T - lymphocytes, NK cells are developmentally closer to T-cells [8]. They secrete a broad spectrum of cytokines that modulate trophoblast behaviour and spiral artery remodelling, thereby playing an essential role in placentation.
Differences between peripheral blood NK cells and uterine NK cells
The first NK cell population described were circulating NK cells in peripheral blood (pbNK), identified in 1975 and named for their ability to lyse leukemic cell lines in vitro [9]. Uterine NK cells (uNK) were classified in the early 1990s; these cells were initially referred to as “large granular lymphocytes of the uterus” based on their size and prominent cytoplasmic granules. Due to shared characteristics with previously described pbNK cells, they were subsequently named as NK cells. When similar cells were later identified in the decidua of certain rodents and primates and confirmed as belonging to the NK lineage, the term uterine NK cells was adopted [10]. Despite the common nomenclature, the antigenic profile, cytokine secretion capacity, and overall function of NK cells are tissue-specific. The primary role of pbNK cells is the control of bacterial and viral infections and tumor surveillance. In contrast, the main functions of uterine NK cells, sometimes further subdivided into NK cells of the proliferative phase (eNK) and decidual NK cells (dNK) [11], include regulation of implantation, placentation, and immunological tolerance of the fetus. During the proliferative phase of the menstrual cycle, uNK cells constitute approximately 20% of all endometrial leukocytes, with their concentration increasing after ovulation [12]. The precise mechanism underlying this increase remains unclear. Possible explanations include local self-renewal of uNK cells (pre-uNK) within the endometrium under the influence of cytokines such as TNFb1 [13], differentiation from bone marrow-derived precursors [14], or recruitment from bone marrow via hematogenous routes or from lymph nodes via lymphatic pathways. Current evidence suggests that migration of precursors through blood or lymphatic circulation is the more likely scenario [15]. Subsequent proliferation of uNK cells within the target tissue is regulated by steroid hormones and interleukins, including IL-12, IL-15, and IL-18. During decidual transformation, NK cells account for approximately 70% of all lymphocytes [16]. This proportion continues to rise during early pregnancy, peaking at the end of the 1st trimester, followed by a gradual decline throughout the remainder of gestation. NK cells lack progesterone receptors on their surface; therefore, the effect of progesterone on NK cell proliferation is indirect, mediated through IL-15 produced by endometrial stromal cells under progesterone influence. Although absolute cell counts vary among individuals, it remains unclear whether tissue concentration alone has a clinically significant impact on pregnancy outcomes [17].
Immunostaining
Immunohistochemistry in the 1980s enabled a more precise definition of individual subpopulations based on the detection of surface proteins. A marker expressed across all leukocyte lineages (T-cells, B-cells, NK cells, monocytes, macrophages, granulocytes) is CD45. The defining features of the NK cell subset are the absence of CD3 and the presence of CD56 on the cell surface (i.e. CD45+CD3–CD56+).
Subsequent classification is based on the expression levels of CD56 and CD16, where CD16 is a membrane glycoprotein belonging to the family of Fc receptors for immunoglobulin G (IgG). CD56^bright NK cells, characterized by high CD56 expression and low CD16 expression (CD56++CD16–), produce large amounts of cytokines and play a regulatory and supportive role in immune responses. They represent the predominant NK cell subpopulation in the uterus and are commonly designated as CD45+CD3–CD56++CD16–. In contrast, CD56^dim NK cells exhibit low CD56 expression and high CD16 expression (CD45+CD3–CD56+CD16+). CD56^dim cells possess strong cytotoxic activity and constitute the major NK cell population in peripheral blood.
Flow cytometry and uNK subpopulations in the endometrium
Until recently, peripheral blood analysis was the primary laboratory approach in reproductive immunology diagnostics. Peripheral blood NK cell levels were considered an important marker, and deviations were interpreted as pathological in relation to implantation failure [17]. After it was demonstrated that NK cells in blood differ in phenotype and function from uterine NK (uNK) cells, attention shifted to endometrial tissue samples obtained by biopsy, particularly during the so-called implantation window. Relatively few studies have focused on the proliferative phase endometrium, while increasing attention is now given to NK cells in menstrual blood (mNK) [18]. Samples from pregnant women are typically obtained from tissue associated with miscarried embryos, raising concerns about potential parameter changes during or after miscarriage, which may affect results.
The most commonly used method for analyzing endometrial samples is flow cytometry, due to its reproducibility, partial automation, and lower interlaboratory variability compared to immunohistochemistry [19]. However, proper pre-analytical handling is crucial, particularly regarding sample preservation and timely processing. For sample analysis, the most frequently used immunofluorescent antibody panel includes anti-CD3, anti-CD56, anti-CD16, anti-CD4, anti-CD8, and anti-CD45. Cells expressing CD45, also known as leukocyte common antigen (LCA), are classified as leukocytes. Combined with SSC-A (Side Scatter Area – a flow cytometry parameter providing information on cellular internal complexity), this allows isolation of lymphocytes. In the next step, staining for CD3 and CD56 enables differentiation of four lymphocyte subpopulations (Tab. 1).
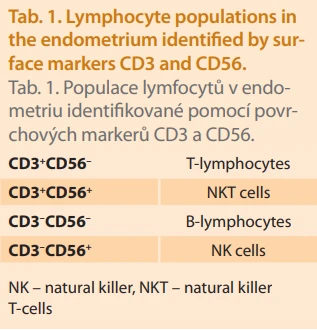
The final step involves stratifying NK cells according to CD56 and CD16 expression levels, providing both absolute and relative counts of NK cell subpopulations (Tab. 2).

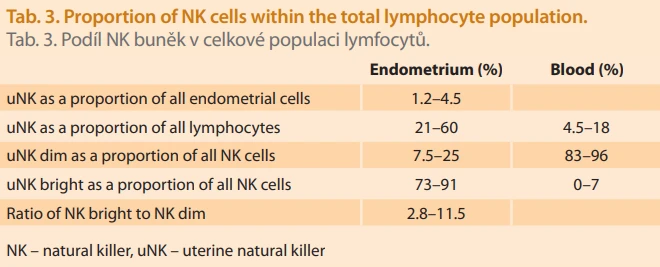
Reference ranges for uterine NK cells and their subsets differ from those of peripheral NK cells and depend on the analytical methodology. Specific percentage ranges for individual subsets vary by laboratory, which often uses internally validated limits [20]. Values may be reported as the proportion of uNK cells out of all cells present in the endometrium. Using this approach, the reference range falls into low values; for example, in the study by Chen et al., it is 1.2–4.5% during the implantation window [21]. Values below or above this interval are considered pathological. However, it can be argued that this assessment of uNK cells does not take into account which specific uNK subpopulations are being evaluated, making the interpretation overly simplified. Therefore, a more detailed classification of individual lymphocyte types within the tissue is used. More commonly, NK cell proportions are evaluated as a percentage of total lymphocytes, followed by regulatory (bright) and cytotoxic (dim) NK cells expressed as a proportion of all NK cells. The Tab. 3 presents the values used by the diagnostic laboratory Gennet Prague, which are internally validated and consistent with published data [22].
Surface receptors of NK cells
CD56 and CD16 are not the only proteins expressed on the surface of NK cells. A particularly important transmembrane structure is the killer-cell immunoglobulin-like receptor (KIR). KIR is a glycoprotein that plays a critical role in NK cell function. KIR receptors occur in two forms: activating KIR and inhibitory KIR. The spectrum and expression levels of individual KIR receptors are independent of the menstrual cycle phase and differ between uterine NK (uNK) cells and peripheral blood NK (pbNK) cells [23]. Another group of receptors under investigation includes the Leukocyte Immunoglobulin-like Receptors (LIR) and Natural Killer Group Receptors (NKG). Each NK cell expresses a unique combination and density of these receptors, resulting in a wide diversity of NK cell phenotypes. For decidual NK (dNK) cells alone, more than 4,000 phenotypes have been identified based on receptor composition. Among these, the most frequent phenotypes occur in approximately 25–45% of individuals, with a predominance of phenotypes carrying KIR receptors [23]. Based on surface receptor profiles, three major subsets of decidual NK cells have been described in the literature: dNK1, dNK2, and dNK3 [24]. Their functional roles and cytokine production vary across different stages of gestation [25]. Peak cytokine production is observed between the 8th and 14th weeks of gestation, coinciding with maximal placental development [26].
When uNK cells encounter extravillous trophoblast (EVT) cells expressing embryonic antigens, a cascade of NK cell-mediated reactions is triggered, which can exert both beneficial and detrimental effects on implantation and embryonic development [27]. Earlier theories suggested that pathological outcomes were caused by activation and cytotoxic action of uNK cells against the trophoblast, leading to its destruction. Current consensus, however, indicates that the presence and activity of uNK cells are essential for implantation and placentation. Their intrinsic cytotoxic activity (CD56^++CD16^−) is low, and uNK cells acquire cytotoxic potential only under specific conditions, such as inflammatory changes in the decidua [28].
Balance between insufficient and excessive trophoblast invasion
As previously noted, the regulation of trophoblast invasion into the decidua is mediated primarily, though not exclusively, through KIR receptors. Extravillous trophoblast (EVT) cells express human leukocyte antigens (HLA) belonging to MHC class I, specifically HLA-C (but not HLA-A or HLA-B, which are also class I antigens but absent on trophoblast), as well as HLA-E, HLA-F, and HLA-G [29]. Trophoblast cells do not express MHC class II antigens (HLA-DP, HLA-DQ, HLA-DR).
The interaction between maternal uNK cell KIR receptors and fetal EVT HLA-C antigens is a major focus of research in the context of assisted reproduction. Maternal KIR receptors interact with HLA-C antigens on EVT, which are of mixed origin: approximately 50% paternal (from the partner or sperm donor) and 50% maternal. In cases involving donor oocytes, the entire HLA-C antigen set on EVT is allogeneic (derived from both the oocyte donor and sperm donor). Both KIR and HLA-C exhibit high polymorphism. Binding of an HLA antigen to a KIR receptor initiates either an activating, or inhibitory signalling pathway, depending on the specific receptor–ligand combination, ultimately ensuring appropriate trophoblast proliferation, invasion, and angiogenesis within the decidua. Controlled activation of uNK cells during this process is essential. Insufficient activation and inadequate trophoblast invasion can lead to placentation disorders. Pathologies arising from these mechanisms include implantation failure, recurrent miscarriage, preeclampsia, and fetal growth restriction. These processes have been well characterized in murine models, which demonstrated the influence of MHC molecules on uNK activation or inhibition, trophoblast invasion, and spiral artery remodelling [30]. Conversely, a widely cited Danish study found no significant association between KIR/HLA-C combinations and preeclampsia incidence [31].
KIR, HLA-C, and regulation of trophoblast migration
As outlined above, the composition of uNK cell receptors is unique to each woman, as is the repertoire of MHC molecules expressed on the trophoblast surface. Focusing on the KIR receptor family, these can be classified into two major genotypes: KIR AA and KIR Bx.
Genetic studies in large pregnancy cohorts suggest that women with the KIR AA genotype have an increased risk of pregnancy complications when the fetus carries an HLA-C allele with the C2 epitope. This combination results in strong inhibition of uNK activity through binding of fetal HLA-C2 to the maternal inhibitory receptor KIR2DL1, and such inhibition of trophoblast invasion has been associated with higher rates of miscarriage, preeclampsia, and fetal growth restriction – although not all studies are in agreement [27]. Conversely, maternal KIR-fetal HLA-C combinations that promote activation via KIR2DS1 or KIR2DS5 are associated with protection against preeclampsia and increased birth weight [32].
Therapeutic interventions and modalities
The increasing availability of diagnostic procedures, such as endometrial immune cell assessment, immunological testing performed on peripheral blood, and, more recently, genetic testing aimed at determining KIR and HLA haplotypes, has led to their overuse. This applies particularly to patients diagnosed with so-called recurrent implantation failure, recurrent pregnancy loss, or unexplained infertility, despite the fact that these diagnostic categories remain poorly defined [33]. Patients carrying these diagnoses are often not only physically but, more importantly, psychologically affected. The innovative nature of assisted reproductive technology, combined with the treatment of this patient population, has contributed to the use of diagnostic and therapeutic approaches that lack support from evidence-based medicine or are used in an off-label manner. Publications addressing this topic, despite occasionally promising results, are typically limited in scope or consist of case studies only [34]. In some instances, immunointerventions have been associated with an increase in surrogate outcomes, such as clinical pregnancy, yet without a corresponding rise in live-birth rates. Reports describing the effects of systemic granulocyte colony-stimulating factor (G-CSF) administration in patients carrying the KIR AA haplotype have shown encouraging outcomes regarding ongoing pregnancy and live-birth rates [35]. However, none of the immunomodulatory interventions are recommended for routine clinical use by professional societies [36–38].
Conclusion
This review provides an up to date perspective on the immunological background of implantation. It outlines the shift in understanding of the role of uterine NK cells in embryo implantation: whereas uNK cells were originally considered potentially detrimental, their function is now regarded as essential for successful implantation. It is evident that uNK cells and their receptors participate in a highly complex process; however, future research may reveal that uNK cells are not the principal regulators. A notable limitation is that most studies investigating uNK cell function rely on decidual tissue obtained from terminated pregnancies or miscarriages. uNK cells in such samples have been directly exposed to EVT or soluble HLA-G, which may have influenced their functional properties and thereby affected study outcomes [26].
Research using uNK cells isolated from the endometrium of non-pregnant women remains relatively scarce, although several studies have reported increased numbers of uNK cells in the luteal-phase endometrium of women with recurrent early pregnancy loss, as well as elevated cytokine expression in women with recurrent miscarriage compared with fertile controls [39]. The use of menstrual blood as an alternative source of endometrial NK cells may broaden future research possibilities [18]. It is also necessary to distinguish between dNK groups 1–3 and clarify their respective functional significance. Given the differences between adaptive and innate immunity, it will be of particular interest to determine whether uNK cell characteristics differ between a first pregnancy and subsequent pregnancies.
A key limitation of the current diagnostic and therapeutic approach to uNK cells lies in the lack of reliable clinical evidence on one hand, and the restricted therapeutic options on the other.
Labels
Paediatric gynaecology Gynaecology and obstetrics Reproduction medicineArticle was published in
Czech Gynaecology
2026 Issue 3
Most read in this issue
- Amniotic band syndrome – a case report of prenatally detected fetal upper limb amputation
- Junctional zone thickness on magnetic resonance imaging – a non-invasive diagnostic method for adenomyosis and beyond
- Perinatal palliative care in a patient with antenatal fetal death due to severe preeclampsia with placental abruption
- Ethanol sclerotherapy for ovarian endometrioma – a systematic review and narrative synthesis